Строение и классификация нейронов
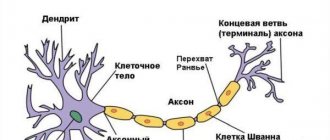
Мозг состоит из миллиардов нервных клеток, или нейронов. Нейрон состоит из трех основных частей: тело нейрона (сома); дендриты – короткие отростки, которые получают сообщения от других нейронов; аксон – длинное отдельное волокно, которое передает сообщения от сомы к дендритам других нейронов или тканям тела, мышцам. Передача возбуждения от аксона одного нейрона к дендритам другого называется нейропередачей или нейротрансмиссией. Существует большое многообразие нейронов ЦНС. Чаще всего классификация нейронов осуществляется по трем признакам – морфологическим, функциональным и биохимическим.
Морфологическая классификация нейронов учитывает количество отростков у нейронов и подразделяет все нейроны на три типа – униполярные, биполярные и мультиполярные.
Униполярные нейроны имеют один отросток. В нервной системе человека и других млекопитающих нейроны этого типа встречаются редко. Биполярные нейроны имеют два отростка – аксон и дендрит, обычно отходящие от противоположных полюсов клетки. В нервной системе человека собственно биполярные нейроны встречаются в основном в периферических частях зрительной, слуховой и обонятельной систем. Существует разновидность биполярных нейронов – так называемые псевдоуниполярные, или ложно-униполярные нейроны.
У них оба клеточных отростка (аксон и дендрит) отходят от тела клетки в виде единого выроста, который далее Т-образно делится на дендрит и аксон. Мультиполярные нейроны имеют один аксон и много (2 и более) дендритов. Они наиболее распространены в нервной системе человека. По форме описано до 60 – 80 разновидностей веретенообразных, звездчатых, корзинчатых, грушевидных и пирамидных клеток.
Классификация нейронов
С точки зрения локализации нейронов, они делятся на центральные (в спинном и головном мозге) и периферические (находящиеся за пределами ЦНС, нейроны вегетативных ганглиев и метасимпатического отдела вегетативной нервной системы).
Функциональная классификация нейронов разделяет их по характеру выполняемой ими функции (в соответствии с их местом в рефлекторной дуге) на три типа: афферентные (чувствительные), эфферентные (двигательные) и ассоциативные.
1. Афферентные нейроны (синонимы – чувствительные, рецепторные, центростремительные), как правило, являются ложноуниполярными нервными клетками. Тела этих нейронов располагаются не в ЦНС, а в спинномозговых или чувствительных узлах черепномозговых нервов. Один из отростков, отходящий от тела нервной клетки, следует на периферию, к тому пли иному органу и заканчивается там сенсорным рецептором, который способен трансформировать энергию внешнего стимула (раздражения) в нервный импульс. Второй отросток направляется в ЦНС (спинной мозг) в составе задних корешков спинномозговых нервов или соответствующих чувствительных волокон черепномозговых нервов. Как правило, афферентные нейроны имеют небольшие размеры и хорошо разветвленный на периферии дендрит. Функции афферентных нейронов тесно связаны с функциями сенсорных рецепторов. Таким образом, афферентные нейроны генерируют нервные импульсы под влиянием изменений внешней или внутренней среды
Часть нейронов, принимающих участие в обработке сенсорной информации, которые можно рассматривать как афферентные нейроны высших отделов мозга, принято делить в зависимости от чувствительности к действию раздражителей на моносенсорные, бисенсорные и полисенсорные.
Моносенсорные нейроны располагаются чаще в первичных проекционных зонах коры и реагируют только на сигналы своей сенсорности. Моносенсорные нейроны подразделяют функционально по их чувствительности к разным качествам одного раздражителя на мономодальные, бимодальные и полимодальные.
Бисенсорные нейроны чаще располагаются во вторичных зонах коры какого-либо анализатора и могут реагировать на сигналы как своей, так и другой сенсорности. Например, нейроны вторичной зоны зрительной области коры больших полушарий головного мозга реагируют на зрительные и слуховые раздражения. Полисенсорные нейроны – это чаще всего нейроны ассоциативных зон мозга, они способны реагировать на раздражение разных сенсорных систем.
2. Эфферентные нейроны (двигательные, моторные, секреторные, центробежные, сердечные, сосудодвигательные и пр.) предназначены для передачи информации от ЦНС на периферию, к рабочим органам. По своему строению эфферентные нейроны – это мультиполярные нейроны, аксоны которых продолжаются в виде соматических или вегетативных нервных волокон (периферических нервов) к соответствующим рабочим органам, в том числе к скелетным и гладким мышцам, а также к многочисленным железам. Основной особенностью эфферентных нейронов является наличие длинного аксона, обладающего большой скоростью проведения возбуждения.
3. Вставочные нейроны (интернейроны, ассоциативные, осуществляют передачу нервного импульса афферентного (чувствительного) нейрона на эфферентный (двигательный) нейрон. Вставочные нейроны располагаются в пределах серого вещества ЦНС. По своему строению это мультиполярные нейроны. Считается, что в функциональном отношении это наиболее важные нейроны ЦНС, так как на их долю приходится 97 %, а по некоторым данным, – даже 99,98 % от общего числа нейронов ЦНС. Область влияния вставочных нейронов определяется их строением, в том числе длиной аксона и числом коллатералей. По своей функции они могут быть возбуждающими или тормозными. При этом возбуждающие нейроны могут не только передавать информацию с одного нейрона на другой, но и модифицировать передачу возбуждения, в частности, усиливать ее эффективность.
Биохимическая классификация нейронов основана на химических особенностях нейромедиаторов, используемых нейронами в синаптической передаче нервных импульсов. Выделяют много различных групп нейронов, в частности, холинергические (медиатор – ацетилхолин), адренергические (медиатор – норадреналин), серотонинергические (медиатор – серотонин), дофаминергические (медиатор – дофамин), ГАМК-ергические (медиатор – гамма-аминомасляная кислота – ГАМК), пуринергические (медиатор – АТФ и его производные), пептидергические (медиаторы – субстанция Р, энкефалины, эндорфины и другие нейропептиды). В некоторых нейронах терминали содержат одновременно два типа нейромедиатора, а также нейромодуляторы.
Другие виды классификаций нейронов. Нервные клетки разных отделов нервной системы могут быть активными вне воздействия, т. е. обладают свойством автоматии. Их называют фоновоактивными нейронами. Другие нейроны проявляют импульсную активность только в ответ на какое-либо раздражение, т. е. они не обладают фоновой активностью.
Некоторые нейроны, по причине их особой значимости в деятельности мозга, получили дополнительные названия по имени исследователя, впервые их описавшего. Среди них пирамидные клетки Беца, локализованные в новой коре большого мозга; грушевидные клетки Пуркинье, клетки Гольджи, клетки Лугано (в составе коры мозжечка); тормозные клетки Реншоу (спинной мозг) и ряд других нейронов.
Среди сенсорных нейронов выделяют особую группу, которые получили название нейронов-детекторов. Нейроны-детекторы – это высокоспециализированные нейроны коры и подкорковых образований, способные избирательно реагировать на определенный признак сенсорного сигнала, имеющий поведенческое значение. Такие клетки выделяют в сложном раздражителе его отдельные признаки, что является необходимым этапом для опознания образов. При этом информация об отдельных параметрах стимула кодируется нейроном-детектором в виде потенциалов действия.
В настоящее время нейроны-детекторы выявлены во многих сенсорных системах человека и животных. Начальные этапы их изучения относятся к 60-м годам, когда были впервые идентифицированы ориентационные и дирекционные нейроны в сетчатке лягушки, в зрительной коре кошки, а также в зрительной системе человека (за открытие феномена ориентационной избирательности нейронов зрительной коры кошки Д. Хьюбел и Т. Визел в 1981 г. были удостоены Нобелевской премии). Явление ориентационной чувствительности заключается в том, что нейрон-детектор дает максимальный по частоте и числу импульсов разряд только при определенном положении в рецептивном поле световой полоски или решетки; при другой ориентации полоски, или решетки, клетка не реагирует или отвечает слабо. Это означает, что имеет место острая настройка нейрона-детектора на потенциалы действия, отражающие соответствующий признак предмета.
Дирекционные нейроны реагируют только на определенное направление движения стимула (при определенной скорости движения). Помимо ориентационных и дирекционных нейронов в зрительной системе обнаружены детекторы сложных физических явлений, встречающихся в жизни (движущаяся тень человека, циклические движения рук), детекторы приближения-удаления объектов. В новой коре, в базальных ганглиях, в таламусе обнаружены нейроны особо чувствительные к стимулам, сходным с человеческим лицом или какими-то его частями. Ответы этих нейронов регистрируются при любом расположении, размере, цвете «лицевого раздражителя». В зрительной системе выявлены нейроны с возрастающей способностью к обобщению отдельных признаков объектов, а также полимодальные нейроны, обладающие способностью реагировать на стимулы разных сенсорных модальностей (зрительно-слуховые, зрительно-соматосенсорные и т. д.).
Нейрон человека[править | править вики-текст]
Основной источник раздела:
Цитология и общая гистология
[2]
Нейроны
(
нейроциты
, собственно
нервные клетки
) — клетки различных размеров (которые варьируют от самых мелких в организме — у нейронов с диаметром тела 4-5 мкм — до наиболее крупных с диаметром тела около 140 мкм). Их общее количество в нервной системе человека превышает 100 млрд. (1011), а по некоторым оценкам достигает одного триллиона (1012).
На сегодняшний день получены неоспоримые доказательства нейрогенеза (процесс регенерации в нервной системе) в субвентрикулярной зоне и субгранулярной зоне (части зубчатой извилины гиппокампа) у млекопитающих, в том числе у людей.[3]
Гибель нейронов в физиологических условиях у взрослого человека
сравнительно невелика и осуществляется механизмом апоптоза. Избыточной потере нейронов препятствует их относительно высокая устойчивость к развитию апоптоза. Гибель нейронов значительно ускоряется в старости, приводя к потере 20-40% клеток в некоторых участках головного мозга.
Гибель нейронов при дегенеративных заболеваниях нервной системы
(болезнях Альцгеймера, Гентингтона, Крейцфельда-Якоба, паркинсонизме, боковом амиотрофическом склерозе и др.) осуществляется вследствие ненормально высокой активности апоптоза, что приводит к резкому снижению их содержания в определенных участках центральной нервной системы (ЦНС). Развитие неврологических нарушений, которые выявляются у 90% больных СПИДом, связано с потерей 40-50% нейронов в коре головного мозга, которые также погибают путем апоптоза.
Функциональная морфология нейрона[править | править вики-текст]
Строение мультиполярного нейрона (по Rohen J.W., Lutjen-Drecoll E. 1982). ПК — перикарион, Я — ядро с ядрышком, ХС — хроматофильная субстанция, НФ — нейрофибриллы (агрегаты элементов цитосклета), Д — дендриты. А — аксон, НСА — начальный сегмент аксона, АХ — аксонный холмик, КА — коллатерали аксона, МО — миелиновая оболочка, УП — узловые перехваты, МБ — моторная бляшка (двигательное нервное окончание на волокне поперечнополосатой мышцы). Синапсы (С): АДС — аксо-дендритический, АСС — аксо-соматический, ААС — аксо-аксональный. Ультраструктурная организация нейрона. Я — ядро (ядрышко показано стрелкой), ХС — хроматофильная субстанция, ЭЦС — элементы цитоскелета (нейротрубочки, нейрофиламенты), МТХ — митохондрии, КГ — комплекс Гольджи, Л — лизосомы, Д — дендриты, А — аксон, АХ — аксонный холмик.
Нейрон состоит из клеточного тела (перикариона) и отростков, обеспечивающих проведение нервных импульсов — дендритов, приносящих импульсы к телу нейрона, и аксона (нейрита), несущего импульсы от тела нейрона.
Тело нейрона (перикарион)[править | править вики-текст]
Тело нейрона (перикарион) включает ядро и окружающую его цитоплазму (за исключением входящей в состав отростков). Перикарион содержит синтетический аппарат нейрона, а его плазмолемма осуществляет рецепторные функции, так как на ней находятся многочисленные нервные окончания (синапсы), несущие возбуждающие и тормозные сигналы от других нейронов.
Ядро нейрона[править | править вики-текст]
Ядро нейрона — обычно одно, крупное, округлое, светлое, с мелкодисперсным хроматином (преобладанием эухроматина), одним, иногда 2-3 крупными ядрышками. Эти особенности отражают высокую активность процессов транскрипции в ядре нейрона. Около ядрышка в нейронах у лиц женского пола часто выявляется тельце Барра — крупная глыбка хроматина, содержащая конденсированную Х-хромосому (особенно заметна в клетках коры полушарий большого мозга и симпатических нервных узлов).
Цитоплазма нейрона[править | править вики-текст]
Цитоплазма нейрона богата органеллами и окружена плазмолеммой, которая обладает способностью к проведению нервного импульса (распространению деполяризации) вследствие локального тока Na+ в цитоплазму и К+ из нее через потенциал-зависимые мембранные ионные каналы. Плазмолемма содержит Na+-K+ насосы, которые поддерживают необходимые градиенты ионов.
Гранулярная эндоплазматическая сеть[править | править вики-текст]
Гранулярная эндоплазматическая сеть хорошо развита, ее цистерны часто образуют отдельные комплексы из параллельно лежащих уплощенных анастомозирующих элементов, которые на светооптическом уровне при окраске анилиновыми красителями имеют вид базофильных глыбок, в совокупности получивших название хроматофильной субстанции (вещества, или телец Ниссля, тигроидного вещества, тигроида). Характер распределения и размеры комплексов цистерн гранулярной эндоплазматической сети (хроматофильной субстанции) варьируют в отдельных типах нейронов (наиболее крупные обнаруживаются в мотонейронах) и зависят от их функционального состояния. При длительном раздражении или повреждении нейрона комплексы цистерн гранулярной эндоплазматической сети распадаются на отдельные элементы, что на светооптическом уровне проявляется исчезновением телец Ниссля (хроматолиз, тигролиз).
Агранулярная эндоплазматическая сеть[править | править вики-текст]
Агранулярная эндоплазматическая сеть образована трехмерной сетью анастомозирующих цистерн и трубочек, участвующих в синтетических процессах и внутриклеточном транспорте веществ.
Комплекс Гольджи[править | править вики-текст]
Комплекс Гольджи хорошо развит (впервые описан именно в нейронах) и состоит из множественных диктиосом, расположенных обычно вокруг ядра.
Митохондрии[править | править вики-текст]
Митохондрии — очень многочисленны и обеспечивают высокие энергетические потребности нейрона, связанные со значительной активностью синтетических процессов, проведением нервных импульсов, деятельностью ионных насосов. Они обычно имеют палочковидную форму и характеризуются быстрым изнашиванием и обновлением (коротким жизненным циклом).
Лизосомальный аппарат[править | править вики-текст]
Лизосомальный аппарат (аппарат внутриклеточного переваривания) обладает высокой активностью и представлен эндосомами и многочисленными лизосомами различных размеров. Интенсивные процессы аутофагии обеспечивают постоянное обновление компонентов цитоплазмы нейрона. При дефектах некоторых лизосомальных ферментов в цитопламзе нейронов накапливаются непереваренные продукты, что нарушает их функции и вызывает болезни накопления, например, ганглиозидоз (болезнь Тэй-Закса).
Цитоскелет нейронов[править | править вики-текст]
Цитоскелет нейронов хорошо развит и представлен всеми элементами — микротрубочками (нейротрубочками), микрофиламентами и промежуточными филаментами (нейрофиламентами). Они образуют трехмерную опорносократительную сеть, играющую важную роль в поддержании формы этих клеток и, в особенности, их длинного отростка — аксона. Многочисленные промежуточные филаменты нейрофиламенты) связаны друг с другом и с нейротрубочками поперечными мостиками; при фиксации они склеиваются в пучки, которые окрашиваются солями серебра. Такие образования (фактически являющиеся артефактами) на светооптическом уровне описаны под названием нейрофибрилл — нитей толщиной 0.5-3 мкм, образующих сеть в перикарионе. Микротрубочки (нейротрубочки) и микрофиламенты имеют такое же строение, как и в других клетках. Клеточный центр присутствует во всех нейронах, его главная функция — сборка микротрубочек.
Включения в цитоплазме нейрона[править | править вики-текст]
Включения в цитоплазме нейрона представлены липидными каплями, гранулами липофусцина (пигмента старения, или изнашивания, который, однако, выявляется даже в нейронах плодов), (нейро)меланина — в нейронах черной субстанции (substantia nigra) и голубого пятна (locus coeruleus).
Дендриты[править | править вики-текст]
Дендриты проводят импульсы к телу нейрона, получая сигналы от других нейронов через многочисленные межнейронные контакты (аксодендритические синапсы), расположенные на них в области особых цитоплазматических выпячиваний — дендритных шипиков. Во многих шипиках имеется особый шипиковый аппарат, состоящий из 3-4 уплощенных цистерн, разделенных участками плотного вещества. Шипики представляют собой лабильные структуры, которые разрушаются и образуются вновь; их число резко падает при старении, а также при снижении функциональной активности нейронов.
Процессы транспорта в нейроне. AAT — антероградный аксонный транспорт (из тела нейрона по аксону) подразделяется на медленный (скорость — 1-5 мм/сут.) и быстрый (100-500 мм/сут.). PAT — ретроградный аксонный транспорт (из аксона в тело нейрона) осуществляется со скоростью 100-200 мм/сут. ДТ — дендритный транспорт (из тела клетки по дендритам) происходит со скоростью около 70 мм/сут. В большинстве случаев дендриты многочисленны, имеют относительно небольшую длину и сильно ветвятся вблизи тела нейрона. Крупные стволовые дендриты содержат все виды органелл, по мере снижения их диаметра в них исчезают элементы комплекса Гольджи, а цистерны гранулярной эндоплазматической сети сохраняются. Нейротрубочки и нейрофиламенты многочисленны и располагаются параллельными пучками; они обеспечивают дендритный транспорт, который осуществляется из тела клетки вдоль дендритов со скоростью около 3 мм/ч.
Аксон[править | править вики-текст]
Аксон (нейрит) — длинный (у человека от 1 мм до 1.5 м) отросток, по которому нервные импульсы передаются на другие нейроны или клетки рабочих органов (мышц, желез). В крупных нейронах аксон может содержать до 99% объема цитоплазмы. Аксон отходит от не содержащего хроматофильной субстанции утолщенного участка тела нейрона — аксонного холмика, в котором генерируются нервные импульсы; почти на всем протяжении он покрыт глиальной оболочкой. Центральная часть цитоплазмы аксона (аксоплазмы) содержит пучки нейрофиламентов, ориентированных вдоль его длины, ближе к периферии располагаются пучки микротрубочек, цистерны агранулярной эндоплазматической сети, элементы комплекса Гольджи, митохондрии, мембранные пузырьки, сложная сеть микрофиламентов. Тельца Ниссля в аксоне отсутствуют. Аксон может но своему ходу давать ответвления (коллатерали), которые обычно отходят от него под прямым углом. В конечном участке аксон нередко распадается на тонкие веточки (телодендрии). Аксон заканчивается специализированными терминалями (нервными окончаниями) на других нейронах или клетках рабочих органов.
Аксонный транспорт[править | править вики-текст]
Аксонный транспорт (ток) — перемещение по аксону различных веществ и органелл; разделяется на антероградный (прямой — из тела нейрона но аксону) и ретроградный (обратный — из аксона в тело нейрона). Вещества переносятся в цистернах агранулярной эндоплазматической сети и пузырьках, которые перемещаются вдоль аксона благодаря взаимодействию с элементами цитоскелета (главным образом, с микротрубочками посредством связанных с ними сократимых белков — кинезина и динеина) процесс транспорта является Са2+-зависимым.
Антероградный аксонный транспорт включает медленный (скорость — 1-5 мм/сут.), обеспечивающий ток аксоплазмы (переносящий ферменты и элементы цитоскелета), и быстрый (100-500 мм/сут.), осуществляющий перенос различных веществ, цистерн гранулярной эндоплазматической сети, митохондрий, пузырьков, содержащих нейромедиаторы.
Ретроградный аксонный транспорт (100-200 мм/сут.) способствует удалению веществ из области терминалей, возвращению пузырьков, митохондрий.
Предполагается, что за счет аксонного транспорта проникшие в нейрон нейротропные вирусы (герпеса, бешенства, полиомиелита) могут распространяться по нейронным цепям. Феномен транспорта используется для изучения межнейронных связей путем введения маркера в область расположения терминалей или клеточных тел и выявления областей его последующего распространения описанными механизмами.
Классификация нейронов[править | править вики-текст]
Классификация нейронов осуществляется по трем признакам: морфологическим, функциональным и биохимическим.
Морфологическая классификация нейронов[править | править вики-текст]
Морфологическая классификация нейронов учитывает количество их отростков и подразделяет все нейроны на три типа: униполярные, биполярные и мультиполярные.
Морфологическая классификация нейронов. УН — униполярный нейрон, БН — биполярный нейрон ПУН — псевдоуниполярный нейрон, МН — мультиполярный нейрон. ПК — перикарион, А — аксон, Д — дендрит(ы).
Униполярные нейроны[править | править вики-текст]
Униполярные нейроны имеют один отросток. По мнению большинства исследователей, в нервной системе человека и других млекопитающих они не встречаются. Некоторые авторы к таким клеткам все же относят амакринные нейроны сетчатки глаза и межклубочковые нейроны обонятельной луковицы.
Биполярные нейроны[править | править вики-текст]
Биполярные нейроны имеют два отростка — аксон и дендрит, обычно отходящие от противоположных полюсов клетки. В нервной системе человека встречаются редко. К ним относят биполярные клетки сетчатки глаза, спирального и вестибулярного ганглиев.
Псевдоуниполярные нейроны — разновидность биполярных, в них оба клеточных отростка (аксон и дендрит) отходят от тела клетки в виде единого выроста, который далее Т-образно делится. Эти клетки встречаются в спинальных и краниальных ганглиях.
Мультиполярные нейроны[править | править вики-текст]
Мультиполярные нейроны имеют три или большее число отростков: аксон и несколько дендритов. Они наиболее распространены в нервной системе человека. Описано до 80 вариантов этих клеток: веретенообразные, звездчатые, грушевидные, пирамидные, корзинчатые и др. По длине аксона выделяют клетки Гольджи I типа (с длинным аксоном) и клетки Гольджи II типа (с коротким аксоном).
Функциональная классификация нейронов[править | править вики-текст]
Чувствительные (афферентные) нейроны[править | править вики-текст]
Чувствительные (афферентные) нейроны генерируют нервные импульсы под влиянием изменений внешней или внутренней среды.
Двигательные (эфферентные) нейроны[править | править вики-текст]
Двигательные (эфферентные) нейроны передают сигналы на рабочие органы (скелетные мышцы, железы, кровеносные сосуды).
Ассоциативные (вставочные) нейроны (интернейроны)[править | править вики-текст]
Ассоциативные (вставочные) нейроны (интернейроны) осуществляют связи между нейронами и количественно преобладают над нейронами других типов, составляя в нервной системе около 99,98% от общего числа этих клеток.
Биохимическая классификация нейронов[править | править вики-текст]
Биохимическая классификация нейронов основана на химических особенностях нейромедиаторов, используемых нейронами в синаптической передаче нервных импульсов. Выделяют много различных групп нейронов, в частности, холинергические (медиатор — ацетилхолин), адренергические (медиатор — норадреналин), серотонинергические (медиатор — серотонин), дофаминергические (медиатор — дофамин), ГАМК-ергические (медиатор — гамма-аминомасляная кислота, ГАМК), пуринергические (медиатор — АТФ и его производные), пептидергические (медиаторы — субстанция Р, энкефалины, эндорфины, вазоактивный интестинальный пептид, холецистокинин, нейротензин, бомбезин и другие нейропептиды). В некоторых нейронах терминали содержат одновременно два типа нейромедиатора.
Распределение нейронов, использующих различные медиаторы, в нервной системе неравномерно. Нарушение выработки некоторых медиаторов в отдельных структурах мозга связывают с патогенезом ряда нервно-психических заболеваний. Так, содержание дофамина снижено при паркинсонизме и повышено при шизофрении, снижение уровней норадреналина и серотонина типично для депрессивных состояний, а их повышение — для маниакальных.
Эфферентные нейроны
Это нейроны, передающие информацию от нервного центра к исполнительным органам.
Пирамидные клетки двигательной зоны коры больших полушарий, посылающие импульсы к мотонейронам передних рогов спинного мозга.
Мотонейроны – аксоны выходят за пределы ЦНС и заканчиваются синапсом на эффекторных структурах.
Терминальная часть аксона ветвится, но есть ответвления и вначале аксона – аксонные коллатерали.
Место перехода тела мотонейрона в аксон – аксонный холмик – наиболее возбудимый участок. Здесь генерируется ПД, затем распространяется по аксону.
На теле нейрона огромное количество синапсов. Если синапс образован аксоном возбуждающего интернейрона, то при действии медиатора на постсинаптической мембране возникает деполяризация или ВПСП (возбуждающий постсинаптический потенциал).
Если синапс образован аксоном тормозной клетки, то при действии медиатора на постсинаптической мембране возникает гиперполяризация или ТПСП. Алгебраическая сумма ВПСП и ТПСП на теле нервной клетке проявляется в возникновении ПД в аксонном холмике.
Ритмическая активность мотонейронов в нормальных условиях 10 импульсов в секунду, но может возрастать в несколько раз.
Проведение возбуждения.
ПД распространяется за счет местных токов ионов, возникающих между возбужденным и невозбужденным участками мембраны.
Так как ПД генерируется без затрат энергии, то нерв обладает самой низкой утомляемостью.
Объединения нейронов
Существуют разные термины, обозначающие объединения нейронов.
Нервный центр – комплекс нейронов в одном или разных местах ЦНС (например, дыхательный центр).
Нейронные цепи – последовательно соединенные нейроны, выполняющие определенную задачу (с этой точки зрения рефлекторная дуга – тоже нейронные цепи).
Нейронные сети – более обширное понятие, т.к. помимо последовательных цепей имеются параллельные цепи нейронов, а также связи между ними. Нейронные сети – это структуры, выполняющие сложные задачи (например, задачи по обработке информации).
Классификация нейронов
I – Морфологическая классификация – по числу отростков и форме перикариона:
А). псевдоуниполярные (с одним отростком) нейроциты, присутствующие, например, в сенсорном ядре тройничного нерва в среднем мозге; псевдоуниполярные клетки, сгруппированные вблизи спинного мозга в межпозвоночных ганглиях;
Б). биполярные (имеют один аксон и один дендрит), расположенные в специализированных сенсорных органах — сетчатке глаза, обонятельном эпителии и луковице, слуховом и вестибулярном ганглиях;
В). мультиполярные (имеют один аксон и несколько дендритов), преобладающие в ЦНС.
II – Функциональная – в зависимости от хар-ра выполняемой клеткой функции (по положению в рефлекторной дуге):
А). Афферентные нейроны
(чувствительный, сенсорный, рецепторный или центростремительный).
К нейронам данного типа относятся первичные клетки органов чувств и псевдоуниполярные клетки, у которых дендриты имеют свободные окончания.
Б).
Эфферентные нейроны(эффекторный, двигательный, моторный или центробежный). К нейронам данного типа относятся конечные нейроны — ультиматные и предпоследние — не ультиматные.
В). Ассоциативные нейроны
(вставочные или интернейроны) — группа нейронов осуществляет связь между эфферентными и афферентными, их делят на интризитные, комиссуральные и проекционные.
Морфофункциональные зоны нейрона.
Микроскопическое и ультрамикроскопические строение зон перикариона, дендритов и аксона. Органеллы общего и специального значения (хроматофильная субстанция и нейрофибриллы).
Транспортные процессы в цитоплазме нейронов.
Морфо-функционал хар-ка нейрона (по Бодиану):
1 – Дендритная зона – это рецепторная зона нервн клетки, она представлена системой суживающихся к периферии цитоплазматических отростков, несущих на своей поверхности синаптические окончания других нейронов.
2 – Зона перикариона – это тело нейрона или скопление нейроплазмы вокруг ядра, здесь расположены органеллы нейрона: митохондрии, КГ, аЭПС, гЭПС, эл-ты цитоскелета.
3 – Зона аксона – одиночный отросток структурно и функционально адаптированный для проведения нервного импульса от тела нервной клетки.
4 – Телодендрий аксона – раветвленные и различно дифференцированные окончания аксонов, где он распадается на тонкие веточки, ктр оканчиваются на др нейронах или клетках рабочих органов.
Морфология нейрона:
Изучение нервн клетки на свеооптическом уровне привело к обнаружению в ее составе специализированных клеточных органелл, ктр были описаны как вещ-во Ниссля
и
нейрофибриллы
.
Вещ-во Ниссля на светооптическом уровнепри использовании основных красителей имеет вид базофильно окрашенных глыбок различного размера и формы, в совокупности они получили название хроматофильная субстанция или тигроидное вещ-во.
На электрограммах аналогом этой субстанции явл-ся гЭПС, хар-р распределения и размеры комплексов ее цистер определяется функциональным статусом и типом нейронов.
Выявленная аналогия между глыбками базофильного вещ-ва и эл-тами гЭПС привела к заключению, согласно ктр вещ-во Ниссля – это хорошо развитая в нейронах гЭПС.
Нейрофибриллы – это система нитей, выявляемых в нейроне при окраске азотнокислым серебром.
Нити толщиной от 0,5 до 3 мкм, идут неориентировано в перикарионе и достаточно упорядоченно в зоне отростков.
При ЭМ оказалось, что нити – это эл-ты цитоскелета нейрона, представленные микротрубочками, микрофиламентами и промежуточными филаментами.
Следовательно, выявляемые в усл-ях СМ нейрофибриллы – это артефакт (рез-т склеивания фибриллярных структур при фиксации материала с последующих отложением красителя на таких комплексах).
Аксонный транспорт (ток) – перемещение по аксону различных вещ-в и органелл; разделяется на антероградный (прямой) и ретроградный (обратный).
Вещ-ва переносятся в цистернах аЭПС и пузрьках, ктр перемещаются вдоль аксона благодаря взаимодействию с эл-тами цитоскелета (с микротрубочками посредством сократ белко – кинезина и динеина); процесс транспорта явл-ся Са2+-зависимым.
Антероградный аксонный траспорт
включает медленный (Ѵ=1-5 мм/сут), обсепечивающий ток аскоплазмы (переносящий ферменты и эл-ты цитоскелета), и быстрый (100-500 мм/сут), осуществляющ ток различн вещ-в, цистерн гЭПС, митохондрий, пузырьков, содержащих нейромедиаторы.
Ретроградный аксонный транспорт
(100-200 мм/сут) способствует удалению вещ-в из области терминале, возвращению пузырьков и митохондрий.
3.3. Нейроны, классификация и возрастные особенности
Нейроны.
Нервная система образована нервной тканью, в состав которой входят специализированные нервные клетки –
нейроны
и клетки
нейроглии.
Структурной и функциональной единицей нервной системы является нейрон
(рис. 3.3.1).
Рис. 3.3.1 А – строение нейрона, Б – строение нервного волокна (аксона)
Он состоит из тела
(сомы) и отходящих от него отростков:
аксона и дендритов.
Каждая из этих частей нейрона выполняет определенную функцию.
Тело
нейрона покрыто плазматической мембраной и содержит в нейроплазме ядро и все органоиды, характерные для любой животной клетки. Кроме того, в ней имеются и специфические образования – нейрофибриллы.
Нейрофибриллы –
тонкие опорные структуры, проходят в теле в различных направлениях, продолжаются в отростки, располагаясь в них параллельно мембране.
Они поддерживают определенную форму нейрона. Кроме того, они выполняют транспортную функцию, проводя различные химические вещества, синтезирующиеся в теле нейрона (медиаторы, аминокислоты, клеточные белки и др.), к отросткам. Тело
нейрона выполняет
трофическую
(питательную) функцию по отношению к отросткам.
При отделении отростка от тела (при перерезке) отделенная часть через 2–3 дня погибает. Гибель тел нейронов (например, при параличе) приводит к дегенерации отростков.
Аксон
– тонкий длинный отросток, покрытый миелиновой оболочкой. Место отхождения аксона от тела называется
аксонным холмиком
, на протяжении 50–100 микрон он не имеет миелиновой оболочки. Этот участок аксона называется
начальным сегментом
, он обладает более высокой возбудимостью по сравнению с другими участками нейрона.
Функция
аксона – проведение нервных импульсов
от тела нейрона
к другим нейронам или рабочим органам. Аксон, подходя к ним, разветвляется, его конечные разветвления – терминали образуют контакты – синапсы с телом или дендритами других нейронов, или клетками рабочих органов.
Дендриты
–
короткие, толстые ветвящиеся отростки, отходящие в большом количестве от тела нейрона (похожи на ветви дерева).
Тонкие разветвления дендритов имеют на своей поверхности шипики, на которых оканчиваются терминали аксонов сотен и тысяч нейронов. Функция
дендритов – восприятие раздражений или нервных импульсов от других нейронов и проведение их
к телу нейрона.
Величина аксонов и дендритов, степень их ветвления в различных отделах ЦНС различна, наиболее сложное строение имеют нейроны мозжечка и коры головного мозга.
Нейроны, выполняющие одинаковую функцию группируются, образуя ядра
(ядра мозжечка, продолговатого, промежуточного мозга и др.).
Каждое ядро содержит тысячи нейронов, тесно связанных между собой общей функцией. Некоторые нейроны содержат в нейроплазме пигменты, придающие им определенный цвет (красное ядро и черная субстанция в среднем мозге, голубое пятно варолиева моста).
Классификация нейронов.
Нейроны классифицируются по нескольким признакам:
1) по форме тела
– звездчатые, веретенообразные, пирамидные и др.;
2) по локализации –
центральные (расположены в ЦНС) и периферические (расположены вне ЦНС, а в спинномозговых, черепно-мозговых и вегетативных ганглиях, сплетениях, внутри органов);
3) по числу отростков
– униполярные, биполярные и мультиполярные (рис. 3.3.2);
4) по функциональному признаку
– рецепторные, эфферентные, вставочные.
Рис.
Рецепторные
(афферентные, чувствительные) нейроны проводят возбуждение (нервные импульсы) от рецепторов в ЦНС.
Тела этих нейронов расположены в спинальных ганглиях, от тела отходит один отросток, который Т-образно делится на две ветви: аксон и дендрит.
Как работает нейрон
Нейроны обмениваются информацией друг с другом на терминалах аксонов и дендритов. Эти терминалы образуют особую структуру, известную как синапс (греч. «Син» = вместе + «гаптеин» = «сцепляться»). Связь между двумя нейронами начинается с потока электрического импульса, известного как потенциал действия, в одном из нейронов. Этот потенциал действия движется вниз по аксону и достигает синаптического терминала. Здесь он запускает выброс нейротрансмиттеров в синаптической щели, крошечном пространстве между терминалами двух взаимодействующих нейронов. Освободившийся нейромедиатор затем связывается с рецепторами на синаптическом окончании другого нейрона и также индуцирует потенциал действия в этом нейроне. Теперь электрический импульс будет проходить по нейрону.